Teoría cromosómica de Sutton y Boveri
Teoría cromosómica de Sutton y Boveri
Teoría cromosómica de la herencia. Más conocida como
Teoría cromosómica de Sutton y Boveri, enuncia que los alelos mendelianos están localizados en los cromosomas. Esta teoría fue desarrollada independientemente en 1902 por Theodor Boveri y Walter Sutton . Permaneció en controversia hasta 1915, cuando
Thomas Hunt Morgan consiguió que fuera aceptada por el mundo.
Fundamentos
Los rasgos de un nuevo individuo son determinados por
genes específicos presentes
cromosomas heredados del padre y la madre. Los
humanos tienen 100.000 genes aproximadamente en los 46 cromosomas.
Los genes que se localizan en el mismo cromosoma tienden a ser
heredados juntos y por esta razón se conoce como genes ligados. En las
células
somáticas, los cromosomas se presentan como 23 pares homólogos para
formar el número diploide de 46. Hay 22 pares de cromosomas apareados,
los autosomas y un par de cromosomas sexuales.
La teoría cromosómica de Sutton y Boveri enuncia que los alelos mendelianos están localizados en los cromosomas.
Esta teoría fue desarrollada independientemente en 1902 por Theodor Boveri y Walter Sutton. También se denomina, a veces, teoría cromosómica de la herencia.
La teoría permaneció controvertida hasta 1915, cuando Thomas Hunt Morgan consiguió que fuera universalmente aceptada después de sus estudios realizados en Drosophila melanogaster.
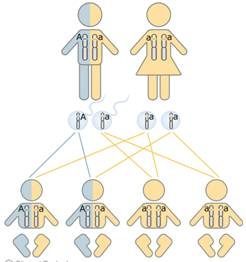
Si el par de cromosomas sexuales es
xx el individuo es genéticamente femenino, si el par es
xy, el individuo es genéticamente masculino.
Un cromosoma de cada par proviene del
gameto materno, el
ovocito, y el otro componente del par proviene del gameto paterno, el
espermatozoide. Así, cada gameto contiene un número haploide de 23 cromosomas y la unión de los gametos en la
fecundación restaura el número diploide de 46.
Las ideas fueron:
La teoría celular, que concibe la célula como la unidad viva autónoma más pequeña; según esta idea, las células son las unidades fundamentales tanto de los organismos unicelulares (como bacterias o protistas), como de los multicelulares, y además constituyen los vehículos de propagación de losorganismos vivos: las esporas, el esperma y los huevos. Un punto muy importante es que la teoría celular considera que las nuevas células proceden de las antiguas, como lo expresó Rudolf Virchow: 'Omnis cellula ex cellula'.
El paradigma mendeliano: según las ideas de Gregor Mendel los elementos heredables se basan en unidades, partículas individuales que pasan de una generación a la siguiente mediante los mecanismos reproductivos. Estas unidades existen en parejas (los alelos), para los cuales pueden existir diferentes variantes. Estas variantes coexisten en los híbridos, y pasan a la generación siguiente en forma de copia simple. Es decir, Mendel establecía el comportamiento combinatorio de los genes (a los que él denominó 'factores') y su segregación en la descendencia.
Los estudios sobre fertilización realizados por Edouard Van Beneden en 1883 demostraron que los pronúcleos masculino y femenino en el zigoto de Ascaris contribuyen cada uno con un juego de cromosomas a la primera división celular. Por otro lado, un influyente citólogo de la época, August Weismann, propuso la cromatina como material hereditario. Incluso estableció la teoría de la línea germinal, y predijo algún tipo de reducción de la información antes de la formación de los gametos. Sin embargo Weismann, en su teoría cromosómica de la herencia de 1892, consideraba que cada cromosoma mitótico contenía el genoma completo de la línea germinal.
Mitosis
Es el proceso por medio del cual una
célula se proviene, dando origen a dos células hija que son genéticamente idénticas a la
célula madre. Cada célula hija recibe el complemento total de 46
cromosomas.
Antes de que una célula entre en
mitosis
cada cromosoma replica su ácido desoxirribonucleico. Durante esta fase
de replicación los cromosomas son extremadamente largos, están dispersos
en forma difusa en el
núcleo y no pueden ser reconocidos con el
microscopio óptico.
Al comienzo de la mitosis, los cromosomas empiezan a enroscarse, a
contraerse y condensarse; estos eventos señalan el principio de la
profase. Cada cromosoma consiste ahora en 2 unidades paralelas, las
cromátidas, que se encuentran unidas en una región estrecha común a
ambas denominada centrómetro.
Durante la profase, los cromosomas continúan condensándose y se
vuelven más cortos y gruesos, pero solo en la prometafase se pueden
distinguir las cromáticas. En el curso de la metafase los cromosomas se
alinean en el plano ecuatorial y entonces resulta claramente visible su
estructura doble.
Cada cromosoma está unido por microtúbulos que se extienden desde
el centrómero hasta el centríolo, formando el uso mitótico. Poco
después, el centrómetro de cada uno de los cromosomas se divide,
señalando el comienzo de la anafase, seguida por la migración de las
cromátidas hacia los polos opuestos del uso.
Por último, durante la telofase los cromosomas se desarrollan y alargan, la envoltura nuclear se reconstituye y el
citoplasma se divide. Cada célula hija recibe la mitad del material cromosómico duplicado.
Meiosis
Es la
división celular que se produce en la
célula germinales para generar los
gametos femeninos y masculinos. En esta se efectúan 2 divisiones celulares sucesivas, la
meiosis I y la meiosis II que reducen el número de
cromosomas a un número haploide de 23.
Las células germinales replican su DNA al comienzo de la primera
división meiótica, de forma tal que cada uno de los 46 cromosomas se
duplica y queda constituido por 2 cromátidas hermanas. Pero, a
diferencia de lo que ocurre en la
mitosis, los
cromosomas
homólogos se aparean alineados entre si mediante un proceso denominado.
El apareamiento es exacto y punto a punto, excepto para la combinación
de X-Y. Luego, los homólogos apareados se separan quedando uno para cada
uno de las 2 células hijas. Poco tiempo después la meiosis II separa
las cromátidas hermanas. Finalmente cada gameto contiene 23 cromosomas.
Entrecruzamiento
Son eventos críticos que se producen durante la
meiosis I y consisten en el intercambio de segmentos de cromátidas entre
cromosomas
homólogos apareados. Los segmentos de cromátidas se rompen y son
intercambiados entre cromosomas homólogos separados. Durante la
separación de los cromosomas homólogos, los sitios de intercambio
permanecen transitoriamente unidos y la estructura cromosómica tiene en
estas circunstancias a la letra X y se denomina quiasma.
En cada meiosis I se producen de 30 a 40 entrecruzamientos, que son más frecuentes entre los
genes localizados distantes entre sí en un
cromosoma.
Fuentes
- Colectivo de autores et Langman. Embriología médica.
Los estudios de Theodor Boveri
Pesar de que en tiempos de
Theodor Boveri se aceptaba que una célula procede de la https://www.blogger.com/blogger.g?blogID=8471508263891909404#editor/target=post;postID=5855798638641481125;onPublishedMenu=overview;onClosedMenu=overview;postNum=2;src=postnamedivisión binaria de una célula madre, no estaba claro cómo la
cromatina (que
August Weismann
denominaba el “plasma germinal”) presente en el núcleo se transmite a
las células hijas de manera que ambas son idénticas a la célula
original, tras la “metamorfosis nuclear” observada por
Walther Flemming, en la cual la masa nuclear se transforma en hebras definidas (los
cromosomas)
que se mueven en el interior celular y luego vuelve a su estado
original. A pesar de que se suponía que dichas hebras transportaban el
material hereditario, el mecanismo permanecía desconocido, hasta que
Boveri demostró que los cromosomas son orgánulos permanentes que se
condensan durante la
mitosis y permanecen difusos durante la
interfase.
Además de establecer la individualidad y la permanencia de los
cromosomas, Boveri dio una descripción moderna del aparato mitótico,
pues fue el primero en identificar los
centrosomas y definir el papel del
huso mitótico
en la distribución de los cromosomas en los polos opuestos de la célula
madre, que darán lugar a las células hijas. Los trabajos de Boveri en
Ascaris
y en embriones de erizos de mar le permitieron observar divisiones
celulares defectuosas, como mitosis multipolares, mitosis monopolares o
medios husos, que después fue capaz de inducir experimentalmente. Esto
le permitió definir tres reglas(1888,1904):
- Los cromosomas durante mitosis son dobles (presentan dos cromátidas),
y cada parte presenta un lateral que se enfrenta hacia un polo del
huso; esta regla implica la idea de que un cromosoma sólo puede
dividirse entre dos células hijas, y la presencia de los cinetocoros, aún no descubiertos, enfrentados en las dos cromátidas para el anclaje de los microtúbulos. Boveri distinguió incluso dos tipos de cromosomas, los que tienen un centrómero localizado (en el erizo de mar) y los que lo tienen difuso (Ascaris).
- Los cromosomas están conectados a ambos polos del huso a través de microtúbulos (MTs).
- Cada cromátida está unida a uno de los dos polos y sólo a uno.
Boveri también identificó que las cromátidas se duplican durante la interfase (1904), y dedujo una correlación muy precisa entre el número cromosómico (la cantidad de cromatina) y el tamaño del núcleo (1905).
De esta forma, para el ciclo cromosómico, Boveri estableció tres
sucesos clave: duplicación de la cromatina durante el periodo de reposo
(interfase), la individualización de las cromátidas durante la
condensación cromosómica y la distribución de las cromátidas en anafase,
una descripción que encaja perfectamente con la visión actual de los
eventos cromosómicos durante el
ciclo celular.
también describió el
centrosoma por vez primera en
Ascaris en 1887,
definiéndolo como un "orgánulo especializado en la división celular".
Boveri identificó claramente el centrosoma como un par de centriolos
rodeados por un material especial, capaz de ensamblar una "esfera de
arquiplasma" que contiene todos esos elementos, que a su vez generan de
forma transitoria una "astrosfera". En 1900, Boveri estableció que los
centrosomas son orgánulos celulares de una única copia. A través de su observación de la dinámica de los cromosomas, llegó a la conclusión de que un
huso mitótico
bipolar típico consiste en realidad de dos medios husos, cada uno
generado por un centrosoma, que se mantienen unidos por el conjunto de
los cromosomas dobles unidos en el extremo de cada áster, de tal manera
que cada cromosoma está unido a ambos polos, y sólo a uno por cromátida.
Por tanto, dedujo que durante la formación de la placa metafásica,
existen fuerzas cromosómicas que parecen contrarrestar la repulsión
existente entre los áster.
Sin embargo, dado que los cromosomas de la línea germinal de Ascaris
son estructuralmene muy polimórficos, Boveri no pudo distinguir la
presencia de cromosomas homólogos en
Parascaris equorum, ni en mitosis ni en el estado de sinapsis meiótica. Fue
Walter Sutton quien reconoció la presencia de cromosomas individuales, identificables
por su tamaño, en espermatocitos de saltamontes, y demostró que 2
cromosomas similares siempre se aparean durante la
meiosis.
Sin embargo, el hecho de que existan cromosomas morfológicamente
distintos no excluye que contengan información genética similar. En
1902, Boveri excluyó esta posibilidad mediante un ingenioso análisis de
dispermia (fertilización por más de un espermatozoide) en erizos de mar,
demostrando que los cromosomas no son equivalentes en el desarrollo
embrionario.
Mediante un análisis cuantitativo publicado en 1907, Boveri estimó el
número de "genóforos" que son esenciales para controlar la ontogenia,
1 que corresponde al número haploide de cromosomas.
A partir de los datos de todos sus estudios citogenéticos, Boveri
llegó a la conclusión de que el aparato meiótico no distingue los
cromosomas homólogos en función de su origen paterno o materno. Por
tanto, esta división puede generar combinaciones múltiples de cromosomas
(2") para crear nuevos juegos haploides en los gametos.
Boveri resumió sus estudios en sus
"Results on the constitution of the chromatic substance in the cell nucleus"
(Resultados sobre la constitución de la sustancia cromática del núcleo
celular), en el que combinaba los hechos observados sobre los cromosomas
con las leyes Mendelianas de la herencia. Como él mismo expresó:"Vemos que dos áreas de estudio que se han desarrollado de manera
independiente han producido resultados que son tan armoniosos como si
uno los hubiera derivado teóricamente del otro".
La confirmación de Thomas Morgan
Thomas Hunt Morgan tenía una formación de biólogo del desarrollo, habiendo recibido un Ph.D. en 1890 en la
Universidad Johns Hopkins por sus estudios en el desarrollo de las arañas marinas, un grupo especializado de
invertebrados,
y en 1891 aceptó un puesto de enseñanza en el Bryn Mawr College. En
1904 la Universidad de Columbia anunció la creación de un nuevo puesto
en zoología experimental, y se lo ofreció a Morgan, quien era amigo de
largo tiempo del director del departamento de zoología, E.B. Wilson.
Wilson convenció a Morgan de que la clave para entender el desarrollo
(esto es, como una célula, el huevo, genera un individuo completo) era
entender la herencia, ya que éste es el medio a través del cual el óvulo
y el espermatozoide transmiten las características de los individuos de
generación en generación.
Morgan inició sus estudios en ratas y ratones, pero éstos se
reproducen tan despacio que no resultaban convenientes para hacer
estudios sobre herencia. Buscando un organismo más apropiado, se decidió
por
Drosophila melanogaster,
la mosca de la fruta, debido a sus características: es un organismo
pequeño (3 mm), fácil de mantener en el laboratorio (se pueden recoger
un millar en una botella de cuarto de litro), es fértil todo el año y
muy prolífica (produce una generación cada 12 días, o 30 generaciones al
año). Además los machos y las hembras se distinguen con facilidad, y el
desarrollo embrionario ocurre en el exterior, lo que facilita el
estudio de las
mutaciones en el desarrollo. Por último,
Drosophila
tiene sólo 4 pares de cromosomas, todo lo cual le convierte en un
organismo muy apropiado para los estudios sobre herencia. Los estudios
de Morgan con
Drosophila comenzaron en 1907. Inicialmente, su
intención era mantener varias generaciones, esperando que apareciera un
mutante ocasional, algo que
Hugo de Vries acababa de observar en plantas. Sin embargo, después de dos años manteniendo las moscas, sus esfuerzos permanecían vanos.
- que los genes deben residir en los cromosomas
- que cada gen debe residir en un cromosoma concreto
- y que el carácter "color de ojos" debe residir en el cromosoma X y
estar ausente en el cromosoma Y, siendo el rojo el color dominante.
Posteriormente, Morgan razonó que los cromosomas son ensamblajes de
genes, puesto que caracteres que se encuentran en un cromosoma
determinado tienden a segregar juntos. Sin embargo, Morgan observó que
esos caracteres "ligados" en ocasiones se separan. A partir de aquí,
Morgan dedujo el concepto de
recombinación
de cromosomas: postuló que dos cromosomas apareados pueden intercambiar
información, e incluso propuso que la frecuencia de recombinación
depende de la distancia entre ambos. Cuanto más cerca estén dos genes en
un cromosoma, mayor será la probabilidad de que se hereden juntos, y
cuanto mayor sea la distancia entre ellos, mayor será la probabilidad de
que se separen debido al proceso de
entrecruzamiento (
crossing-over).
En resumen, Morgan sugirió que la intensidad del ligamiento entre dos
genes depende de la distancia entre ellos en un cromosoma. Basándose en
esas observaciones, un estudiante del grupo de Morgan,
Alfred Henry Sturtevant,
llegó a la conclusión de que las variaciones en la intensidad de
ligamiento podían utilizarse para mapear los genes en los cromosomas,
definiendo la distancia relativa unos de otros: un año después de que
Morgan hubiera identificado la mosca de ojos blancos, Sturtevant
estableció el mapa genético para los genes ligados al sexo. Hoy en día,
el Morgan es la unidad de medida de las distancias a lo largo de los
cromosomas en la mosca, el ratón y en humanos.
Morgan fue galardonado con el
Premio Nobel de Fisiología o Medicina en
1933 por la demostración de que los
cromosomas son portadores de los
genes.